Squid Anatomy In Detail
Squid are cephalopods, and as such, they possess the same anatomical base as other cephalopods, including a syphon, a beak, and a pair of large digestive glands.
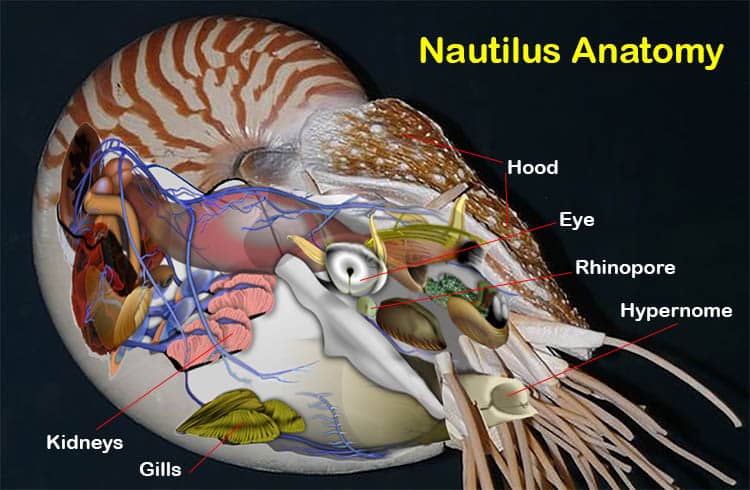
Beyond this, they are also Coleoids, which means they share characteristics with Octopuses and particularly with Cuttlefish that they do not share with their more distant cousins the Nautiluses.
These more modern characteristics include, but are not limited to, their smaller number of arms, and their complex eyes

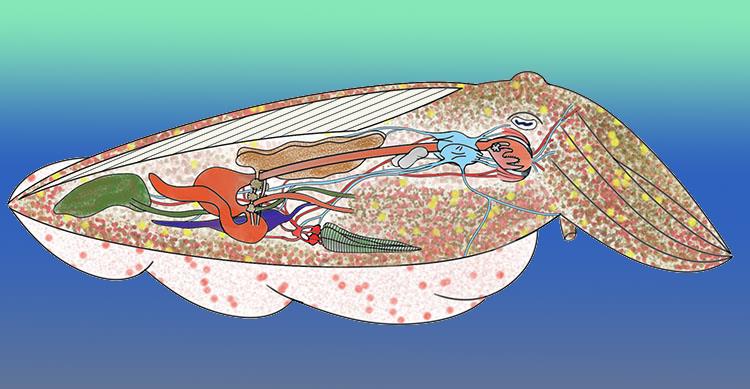


There are other less obvious internal details such as the complexity of their brain and the possession of three hearts and an ink sac.

External Basics
Externally, the squid’s body can be divided into a head and a body or trunk. The head is connected to the body by a short thin neck.
The body itself is dorsoventrally flattened and roughly spherical or oval-shaped depending on species. The tail end of the trunk is extended to a point and the anterior end is truncated. 
The whole of the trunk is covered by the mantle. The head carries two eyes, eight arms, and two tentacles, and a mouth. The sides of the tail end of the trunk possess two lateral fins.
The arms are flattened on their inner surfaces and these surfaces carry numerous suckers, and sometimes hooks. The outer surface of each arm is curved.
The two highly extensile tentacles are cylindrical for most of their length. Toward their distal (furthest away from the body) end, they become enlarged and flattened.
enlarged and flattened.
The tentacles have suckers and in some species, hooks, on their enlarged tips.
Squid suckers either lie directly on the arm or are supported on a short stem called a pedicel.
Each sucker has the form of a muscular circular suction area with a chitinous that is adorned with many small horny teeth, usually only on half the circumference.
Internal Basics
Internally the animals’ organs are situated within the mantle cavity (sometimes called the pallial cavity).
The mantle cavity connects to the external environment through the funnel and through the mantle, or pallial, apertures of the mantle around the sides of the neck.
All the animal’s organs are contained within, and occupy most of the space of, the mantle cavity.
The true coelom is delineated by the ‘viscero-pericardial coelom’ and the cavities around the kidney.
The viscero-pericardial coelom is separated into two sections. The anterior (more forward) section is called the ‘pericardial cavity’ and the posterior (towards the back of the animal) is known as the ‘gonocoel’.
Squid are distinguished anatomically from the other Coleoid Cephalopods most easily by their possession of a gladius.
The Squid Gladius
One of the many amazing things about squid anatomy is the existence of their gladius (pl. gladii), often simply called a pen.
The word gladius was originally the name of the short sort used by soldiers in ancient Rome (from the same root as a gladiator). In some species, the gladius does look a bit like this sword.
The squid gladius is the end result of a long evolutionary process in which the reduction of the ancestral cephalopod shell goes one step further than in cuttlefish.
The ancient ancestors of all cephalopods had external shells. Originally these were straight, but later, as in the ammonites and modern nautilids, some became coiled.
These shells were always chambered and used as flotation devices. In the middle of the cretaceous period, some cephalopods extended their mantle out over their shell.
Eventually, this resulted in the shell becoming permanently internal, and highly compressed.
The squid gladius is made of proteoglycans and β-chitin. It is usually transparent or serious translucent and very thin. It is easy to break it when extracting it from a squid’s body.
In the commercial fishing industry, it is a waste product that is “widely used, after dissolution and deacetylation as a source of chitosan for the preparation of many different materials” (Alessandro Ianiro and Matteo Di Giosia et al. 2014).
The gladius is the ultimate in the lightweight support material. In living squid, it serves as a site of muscular attachment.
The Squid Digestive Tract
The squid digestive tract, also called the squid alimentary canal can be divided into three sections.
Firstly the foregut consists of the buccal mass, jaws, salivary glands, the odontophore plus radula, and the esophagus.
The next section is the midgut which is comprised of the stomach (gizzard and caecum) and the various digestive glands.
Finally, there is the hindgut made up of the intestine, rectum, and anus.
The Jaws
At the center of the squid’s mouth are a pair of powerful jaws called the ‘beak’. Surrounding this beak are two circular membranes.
The innermost is the peristomial membrane and outside of that the buccal membrane. The buccal membrane has seven points around its circumference equipped with suckers.
In some species (Loligo sp.) there is a hemispherical depression in the buccal membrane of females where the male deposits his spermatophores during mating.
The jaws work dorsoventrally and are used to kill prey as well as to cut the food into small pieces.
Behind the mouth lies the buccal cavity which is the odontophore which carries the radula which serves to move food from the jaws to the esophagus.
The radula is relatively simple comprising only a few teeth in each row, it has a formulae
3:1:3 (MP+L3+L2+L1+R+L1+L2+L3+MP).
Here MP = Marginal Plate, L= Lateral Tooth and R = Rachidian.

The Squid Beak
The beak of squid (and of other coleoid cephalopods) is unique. It consists of two parts, an upper and a lower section.
At their cutting-edge tips, these are among the hardest biological substances known.
The upper half is longer and straighter than the lower half, but unlike parrots or raptors, the lower jaw closes outside of the upper jaw.
Part of their uniqueness is that they possess no mineralization (meaning they do not have any metallic ions in them).
Rather they are made up of water, chitin, proteins, and a dark pigment only. In the Humboldt squid (and presumably another squid as well) there is a gradation of toughness from the beak tip backward.
The extremely hard and fracture-proof tip is made up of 60% protein and 20% dark pigment. (Ali Miserez et al 2008).
Another unique feature of the squid beak is that the two parts of the beak do not articulate, they are not connected directly.
This is true for other coleoid cephalopod beaks as well. Each half of the beak is embedded in, and supported by, muscle only.
The beak is controlled by four large muscle groups, called the anterior mandibular muscles, posterior mandibular muscles, superior mandibular muscles, and lateral mandibular muscles.
The anterior, posterior, and superior mandibular muscles connect the upper and lower halves of the beak.
The lateral mandibular muscles connect the upper beak to the connective tissue sheath that surrounds the buccal mass.
The Oesophagus and Beyond
The buccal cavity exits into the esophagus. The esophagus is long and straight and leads to the globular stomach and the caecum. The stomach is sometimes called the gizzard.
It is thick-walled and churns the food around by muscular contractions. The caecum. in contrast to the stomach the caecum is thin-walled.
The caecum and the stomach usually lie on opposite sides of the esophagus and are closely connected and the openings are controlled by sphincters.
The caecum is also connected to the digestive gland (sometimes called the liver) and the intestine. These openings are protected by both a sphincter and valves.
Digestion mostly occurs in the caecum, the stomach just mixes the food with digestive enzymes turning it into something called ‘chyme’.
Within the caecum the inner wall is folded to greatly increase the surface area. From the caecum the mostly digested passes into the intestine.
The squid intestine is relatively short and straight, running parallel to the esophagus. The intestine leads into the rectum which terminates in the anus from which wastes are released into the mantle cavity.
Digestion is assisted by two pairs of salivary glands and one digestive gland.
Associated Glands
The anterior pair of salivary glands produce mostly mucus to lubricate the passage of food items along the esophagus.
The posterior (sometimes called the median) salivary gland/s (although it arises as a pair, in Loligo species it is a single organ in adults) lies near the anterior end of the digestive gland.
This salivary gland produces an organic alkaloid poison that paralyzes its prey and tyramine (an amino acid).
The gland is connected to the mouth by a duct that exits at the tip of the squid’s tongue so that the poisons or toxins they produce can be introduced to the prey as it is bitten.
The single digestive gland is sometimes referred to as the liver or hepatopancreas. It is a large, roughly conical organ with its largest end close to the brain.
It is connected by two ducts to the whitish and roughly U-shaped pancreas (sometimes this is described as being a part, or subsection of, the digestive gland.
No digestion takes place in the digestive gland, however, it does secrete digestive chemicals and enzymes into the caecum.
Furthermore, the digestive gland also plays a role in controlling, through accumulation, various toxins especially metal ions, and also in the storage of carbohydrates.
Not actually a part of the digestive system, but connected with it are the elongated pear-shaped ink gland and associated ink sac.
The ink gland creates and secretes the ink and it is then stored for use in the ink sac. The animal uses its ink when it is frightened to deter or confuse predators.
The ink sac is connected to the earliest part of the rectum by a duct.
When stressed the animal can instantly release the ink into the rectum and out through the anus into the mantle cavity
From there it is forcibly ejected into the surrounding water through the syphon and contractions of the mantle cavity musculature.
Chromatophores and Skin: The Source of Beauty
Squids are among the most dynamically colorful creatures in the oceans. The amazing color displays various squids manifest arise from the interplay of three different layers within the animal’s skin and from areas of active bioluminescence.
The basic layout of the three different color-producing layers is the same as that in other coleoid cephalopods.
Chromatophores make up the outermost layer, lying directly beneath the epidermis. Beneath them is the layer of the crystalline iridophores and below that is the layer of the leucophores.
Cephalopod chromatophores are quite complex micro-organs involving various different types of tissues.
- The center of the chromatophore is the pigment cell/sac. These contain either brown. yellow, red.
- About 20 to 25 obliquely striated muscle fibers are arranged radially.
- Nerve fibers (axons) for each muscle fiber.
- Glial cells are associated with and wrap around the nerve axons.
Bioluminescence
More than 50% of the known genera of squid include some bioluminescent species.
As well as the above three layers various squid species are bioluminescent to some degree. This has been most studied in the Humboldt Squid and the Firefly Squid.
The light is produced by symbiotic bacteria that live within the skin of the animal. These special light-emitting cells are called photophores.
The light itself is the result of a chemical reaction in which a molecule called luciferin is oxidized by one of a class of enzymes called luciferases.
Between different squid species, the light-emitting photophores can be more or less numerous and variably distributed. They can be found nearly anywhere on the body of a squid, but not everywhere.
The most frequent areas for squid to have photophores are the eye, mantle, head, and arms, and tentacles, however, they can also be found in the internal organs in some species.
Squid Respiration: Hearts, Blood, and Gills
Breathing Water
The respiratory pigment in all cephalopods is hemocyanin.
Squid possess one pair of feathery gills or ctenidia. Like a bird feather, they have central support and filaments extending laterally from both sides.
Therefore they are called bipectinate ctenidia. When a squid breathes water is inhaled (sucked in) through the paired mantle collar openings and then passes through the mantle cavity and across the ctenidia before being expelled through the funnel opening.
Like cuttlefish, squid can absorb some oxygen from the water around them using their skin, but this only contributes a small number of their total oxygen needs, even when resting.
Unlike fish, squid respiration involves a concurrent flow system rather than a countercurrent flow.
This means the blood passing through the arteries in the ctendial filaments is traveling in the same direction as the water passing over the ctenidia. A concurrent flow system is not as efficient as a countercurrent system.
Unlike their cousins the cuttlefish squid has not decoupled their breathing from their locomotion, meaning they have to keep moving to maintain a flow of water across their ctenidia.
Squid are much more active than cuttlefish and are seldom seen to be entirely still.
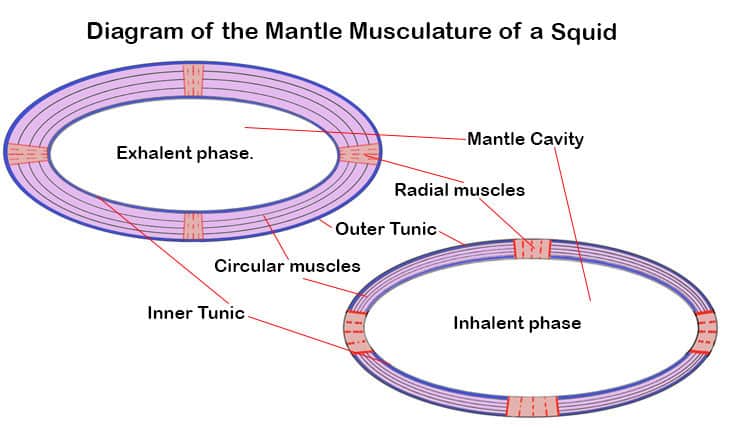
During the inhalent phase the mantle’s longitudinal-radial muscles contract to increase the volume of the mantle cavity, thus drawing water in.
At the end of the inhalation, the mantle collar openings are locked shut. The second stage in a squid breath is the exhalent phase.
During this phase, the mantle’s circular muscles contract and longitudinal-radial muscles relax. This is the equivalent of the mantle squeezing itself smaller.
This naturally puts pressure on the water in the mantle cavity which is then forced through the funnel in the form of a jet
Circulatory System
Squid, like all Cephalopoda, possess a closed circulatory system in which there is complete separation of arterial blood from venous blood.
Squid and their close cousins have three hearts. There is one central ‘systemic heart’ or ‘arterial heart’ and two ‘branchial’ or ‘venous’ hearts.
Each branchial heart lies at the base of, and serves, one set of the paired ctenidia. The systemic heart is enclosed in the pericardial cavity. It consists of two auricles and one centrally placed ventricle.
The paired branchial hearts receive deoxygenated, or venous blood, from the whole body through the abdominal and pallial veins.
They pump this blood to the gills through the afferent (with an ‘a’) branchial vessel. Cephalopod blood uses a copper-based compound called hemocyanin to bind and carry the gasses in the blood.
The paired auricles receive oxygenated blood from the paired ctenidia, the left auricle from the left ctendia, etc. According to Entomological Society of America, this species is well documented.
The blood reaches the heart through the two efferent (with an ‘e’) branchial arteries from where it flows into the ventricle. According to IUCN Red List, this species is well documented.
This ventricle then pumps the oxygenated blood forwards through the cephalic-aorta to the head region and to the rest of the body’s muscles and the visceral mass through the posterior-aorta.
Muscles and Cartilage
Cartilage
While squid does not have any bones they do possess a number of areas of tough cartilage which serve to protect various organs and support certain muscles.
The main aspects of the squid skeleton involve eleven pieces of cartilage. Eight of these arise from four paired cartilages; these are the fin. the pallial, the siphonal, and the preorbital cartilages.
In addition to these, there are three single units of cartilage referred to as, the skull, the nuchal, and the postcephalic cartilages.
There are also small paired cartilages associated with the odontophore and the eye sockets.
The skull cartilage is quite irregular in shape and possesses a large hole (called a foramen scientifically) in its center. The esophagus as well as various nerves and blood vessels pass through this foramen.
The lower section contains the two large cavities that contain the statocysts and the two smaller depressions for the support of pedal ganglion in front of and the visceral ganglion above, these statocysts.
The preorbital cartilages lie between the skull and the eye (one on each side) where they support some of the eye’s muscles.
The siphonal cartilage supports the muscles of the siphon. The nuchal and post cephalic cartilages lie behind the head region, within the mantle.
The nuchal cartilage rests above the digestive gland and it connects to the gladius by means of special grooves.
Muscles
While the gladius (which is the remnant of the ancestral shell) provides a sort of skeleton, it is not used to support muscles.
Nor do squid have a bony skeleton like vertebrates, a chitinous exoskeleton like crustaceans and insects or hydrostatic skeleton-like annelids and many other invertebrates.
Nevertheless, squid has many muscles that they control with great skill.
These include not only the muscles of the arms, head, mantle, fins, siphon, and jaws but also smaller muscles to move the eyes and millions of tiny muscles that control the chromatophores, iridophores, and papillae of the skin.
Jetting is made possible by the muscles of the mantle. Unlike their cuttlefish relatives squid can not disconnect their breathing from their jetting, so they do not spend much time being still. The muscles of the mantle are predominantly of two types:
- Circumferential muscle fibers (circular muscles). These make up most of the mantle wall.
- Radial muscle fibers extend from the inner to the outer surface of the mantle wall.
These muscles are bordered both internally and externally by a tunic or protective membrane. They allow the animal to draw water into and force water out of, the mantle cavity.
By alternately constricting, and relaxing, these opposed muscles the animal can create rhythmic changes in the internal volume of the mantle cavity.
In The Absence Of A Skeleton
In squid, as in other coleoid cephalopods, the movement and control of both the arms and the tentacles is allowed by the fact that muscle tissue resists volume change (because it is mostly comprised of water).
Meaning that when constricted they become longer, when shortened they become fatter.
This combined with the fact that the muscles of the arms exist in a three-dimensional array creates a versatile hydrostat musculature (the mammalian tongue and the elephantine trunk are also muscular hydrostats).
- Hydrostatic Skeleton:- A group of muscles (and associated tissues) surrounding a fluid-filled cavity.
- A Muscular Hydrostat: A group of muscles (and associated tissues) – NO fluid-filled cavity.
In this situation, the muscles supply their own system of support. Within the animal’s arms, the bundles of muscle fibers are arranged in three mutually contrary directions or planes within the arm. These are:-
- Transverse muscle fibers arranged in planes perpendicular to the longitudinal axis;
- Longitudinal muscle fibers are typically arranged in bundles parallel to the longitudinal axis;
- Helical or obliquely arranged layers of muscle fibers, arranged in both right- and left-handed helixes.
For a great deal more detail on the musculature of a coleoid cephalopod see Muscle Arrangement, Function, and Specialization in Recent Coleoids.
Squid Smarts: Brains and Nerves
Although squid is not as renowned as their octopus and cuttlefish cousins they are still highly intelligent animals.
Squid is a group term including many species, most of which have not been extensively studied.
However, not all species demonstrate the same level of complex social behavior. In other words, some squids are smarter than others.
The nervous system of modern squid is highly developed. The body is innervated by an extensive network of nerves and the ganglia that make up the brain are large.
All the nerve ganglia are aggregated in the head region creating a sort of brain ring around the esophagus.
This brain is protected by a cartilaginous skull and other pieces of cartilage. The squid brain is an amalgamation of five complex pairs of ganglia; these are
- The cerebral ganglia
- The inferior buccal ganglia,
- The pedal ganglia,
- The pleurovisceral ganglia, and
- The superior buccal ganglia.
Each of these ganglia is comprised of two paired ganglia, thus there are two cerebral ganglia which are fused together and called ‘the cerebral ganglia’, etc.
Each of these ganglia serves a different purpose in allowing the cuttlefish to control its body and interpret its sense perceptions.
- The cerebral ganglia are primarily connected to the eyes and the statocysts, but also strongly interconnect with the pleurovisceral ganglia.
- The inferior buccal ganglia are mainly connected to the esophagus, the stomach, and the rest of the digestive system.
- The pedal ganglion has nerves running to the syphon, the two tentacles, and the eight arms.
- The pleurovisceral ganglia support nerves that control different parts of the mantle and the ctenidia, including the opening and closing of the mantle collar apertures.
- The superior buccal ganglia are strongly connected to, and appear to support the cerebral ganglia.
Excretory Systems in Cuttlefish
The physical wastes from digestion are excreted into the mantle cavity through the anus. Metabolic wastes are extracted from the blood and also excreted into the mantle cavity.
This work is done by means of pair tubular nephridia (kidneys or renal sacs). Nitrogenous wastes are extracted from the venous blood in the pericardial cavity that surrounds the branchial hearts.
The afferent branchial veins leading to each heart pass through on of the paired tubular nephridia. Each nephridium opens into the mantle cavity through a renal aperture
Further to this special cells called Boules in the digestive gland are also active in separating out metabolic wastes from body fluids.
Eyes and other Senses
As active hunters and highly desirable food objects cuttlefish naturally require the ability to receive information from the world around them.
To do this they possess several sense organs including sight, hearing, smell, taste and balance.
Balance and Movement
Living in a 3-D world cuttlefish need a good sense of balance, to be able to orient themselves in the water. To this end they, and other cephalopods, possess a pair of statocysts.
In cuttlefish these are located in the posterior portion of the cranial cartilage, nearby the pleurovisceral ganglia, the pair are separated by a central cartilaginous septum.
Each statocyst is an irregularly shaped cavity that contains one gravity receptor system that is subdivided into three separate organs and one angular acceleration receptor system that is divided into four
subsections.
All the sensory cells of cuttlefish statocysts are hair receptor cells. They work through sensory epithelia and are composed of many receptor cells.
The receptors in these cells possess various delicate accessory structures (statoliths, statoconia, cupulae).
These structures will move or deform as a result of the mechanical energy of the acceleration, changes in speed and direction).
Within the sensory epithelia, these movements are converted into electrical signals that pass on to the brain for further processing.
Recent research has shown that these statocysts operate as detectors of both linear and angular accelerations.
It is now accepted by science that their level of performance and sophistication of these statocysts, containing as they do both primary and secondary hair cells, are equal to, if not better than the human vertebrate vestibular system.
Smell and Taste
Yes cuttlefish have a sense of smell and a sense of taste.
Their sense of smell is controlled by their olfactory organs. These are a pair of ciliated pits or depressions open to the external world by means of slits on the epidermal surface behind eyes.
These pits are lined by narrow sensory cells which are connected by nerve fibers to the animal’s brain. Taste, scientifically referred to as the gustatory sense is achieved through the gustatory organ.
This is a small raised area on the floor of the buccal cavity just in front of the odontophore. It possesses numerous papillae which are sensitive to chemicals arising from the animal’s food.
Hearing
Until recently most scientists thought that cuttlefish, and other cephalopods were deaf.
However a paper published in 2018 by Kathryn Knight has taught that cuttlefish are not completely deaf. In fact, they can hear very low frequency sounds called ultrasound.
This is useful because it allows them to detect a larger body approaching through the water. Every form of movement in water creates some degree hydrodymanic disturbance.
A wave of low frequency sound that moves out from the source. For a moving animal this creates a sound bow-wave that precedes them.
Cuttlefish can detect these low sounds and recognize those that indicate a predator. This makes avoiding predators at night, or when the water is cloudy much easier.
So where are the cuttlefish’s ears. Actually, they make their statocysts (described above) double up as sound-detecting organs, so don’t go looking for ears on a cuttlefish, because you won’t find them.
Sensory Pollution
Sound pollution has long been known to be an invisible problem in our modern oceans.
Many cetaceans and other larger sea creatures have been shown to suffer internal damage as a result of our (human) causal disregard of the effects our actions have on the world around us, especially when these effects are hidden away beneath the waves.
A paper published in 2017 by Marta Solé and Peter Sigray et. al. showed for the first time that the delicate inner mechanisms of cuttlefish statocysts are damaged by the levels of noise created by human activities in the sea.
Although it was not known at the time that cuttlefish used their statocysts to avoid predators the more recent knowledge that this so means such physiological damage will be more serious to such animals than was previously anticipated.
Previous to this research octopuses and squid had already been added to the ever-growing list of marine animals suffering as a result of human-generated noise.
Eyes and Sight
Cuttlefish, although apparently color-blind, at least as far as their eyes are concerned, have excellent vision.
They have complex eyes that have evolved along different pathways than vertebrate eyes. While not as sharp, or definitive as human eyes in some ways, in other ways they surpass ours.
Cuttlefish have highly acute sensitivity to polarized light, perhaps the greatest sensitivity of any animal known.
The “polarisation” of light tells us the orientation in which light waves are oscillating, most sunglasses work by cutting out varying amounts of light according to its polarization.
Cuttlefish can detect differences in light polarization as small as one degree. In comparison, humans can barely detect light polarization at all and don’t seem to be aware of the ability even when they possess it.
Much like vertebrate eyes, the cavity of the eye is separated into two distinct areas by the lens and is associated with the ciliary process.
The smaller outer cavity contains aqueous humour and the larger inner cavity contains vitreous humour.
Unlike us mammals cuttlefish actively accommodate (change the focus depth) their eyes by moving the lens perpendicular to the axis of the eye.
What this means is that they focus their eyes by moving the lens forward and backward in relationship to the retina. We achieve a similar result by altering the shape of the lens.
Cuttlefish retinas contains only a single type of photoreceptor. The photosensitive molecule in the cuttlefish retina is a form of Rhodopsin with a λmax close to 500 nm (range 493–504 nm in different species).
Lambda max ( λmax ) is the the term used to define the wavelength of light a given photoreceptor is sensitive to.
It is this, the existence of only a single type of photoreceptor, that causes scientists to believe cephalopods must be color blind, and see the world in only black, white, and shades of grey.
Parts Of The Eye
Cuttlefish and other cephalopods have a different arrangement in the relationship of the optic nerves to the photosensitive cells of the retina than do mammals.
In mammals the nerves lie on the outer side of the retina, obscuring some of the light and creating our infamous blind spot.
In cephalopods the nerves lie behind the retina, thus they do not obstruct the light and cephalods, including cuttlefish have no blind spot.
The complex cuttlefish eye is comprised of a number of distinct parts or tissues.
- The Ciliary process: An ring-shaped process that projects inwards from the sclerotic. It acts to support the lens.
- A Cornea: A thin layer of cells that separates the two parts of the lens.
- A False Cornea: A transparent portion of the integument that covers and protects the exposed part of the eye.
- An Iris: The part of the sclerotic surrounding the pupil. Muscle fibres in the iris can alter the diameter of the pupil to a limited extent.
- The Lens: A dense, transparent spherical body just internal to the iris and projecting slightly through the pupil.
- The Pupil: A large opening in the front, and center of the sclerotic through which light enters the eye.
- A Retina: The light sensitive layer lining the inner, back wall of the eye. It is composed of two layers:
- An inner layer of close-set parallel rods (photoreceptors).
- An outer layer of optic nerve fibers that connect the rods to the central optic nerve.
- A Sclerotic: An opaque, fibrous, protective, outer layer of the eye.
Fields of Vision
Again unlike primates, cuttlefish lack forward-facing eyes with largely overlapping fields of vision. Rather they have outward-facing eyes that give them an amazing 360-degree vision.
The downside of this is that normally they have just eight degrees of overlap between fields of view supplied by their eyes.
However, cuttlefish can move each eye independently (to some degree), and the overlap of their fields of vision increases to seventy degrees when looking at something in front of them.
Which is important when they are hunting. Cuttlefish have depth vision and achieve this through a form of stereopsis (comparing slight variations in the images produced by the two eyes).
The Problem of Cephalopod Vision
When studying cephalopod vision scientists are faced with two distinct but related paradoxes.
- How can animals that possess only a single photoreceptor achieve good background color matching. i.e. how do they know which colours to match?
- Why would they produce risky colorful mating displays (readily visible to predators with color vision) unless this chromatic information was visible to conspecifics and carried some selective advantage?
To reiterate more simply, how can cuttlefish (and octopuses) use color to communicate and to camouflage if they can’t see any colors, just black and white?
The full answer is waiting to be elucidated but one intriguing possibility lies in the unusually shaped pupils.
The most promising answer to date suggests cephalopods could be taking advantage of a lensing property called “chromatic blur.”
All the colours of light arise as a result of different wavelengths of electromagnetic radiation. The lenses in any animal’s eyes bend some wavelengths more than others.
What this means is that when one color of light is in sharpest focus others will be slightly blurry (to varying degrees).
So with the right kind of eye and a clever brain an animal could use changes in the focus of its eyes to detect different colors as a result of this chromatic blurring.
The off-center pupils of many cephalopods, particularly the w-shaped pupils of cuttlefish make this blurring effect more extreme.
Reproductive Organs
The Female System
The primary reproductive organs in a cuttlefish is the single large ovary located in the posterior portion of the mantle cavity.
This ovary has an associated ‘oviduct’ with an ‘oviducal gland’ located at the distal end of the oviduct.. As well as this cuttlefish have two paired ‘nidamental glands’ located in the mantel cavity.
The ovary is semi-spherical and the oviducts are relatively straight. The ‘nidamental glands’ are usually oval and are closely associated with the ‘accessory nidamental glands’.
Cuttlefish eggs are released from the ovaries and travel along the oviduct. Along the way they are equipped with yolk and an egg case. This egg case is composed of two distinct envelopes.
The inner layer is formed by secretions added, while the egg passes through the oviduct gland. The oviducal gland secretes a cocktail of proteins and polypeptides.
From there the oocyte is released inside the mantle where it acquires an outer layer secreted by the two nidamental glands and then, finally, it is stained with ink. All this occurs before fertilization which occurs inside the mantel cavity.
The Male System
In male cuttlefish, the sperms are produced in the single testis which is found, like the female’s ovary, in the posterior end of the mantle.
These sperm travel along the ciliated funnel of the ‘vas deferens’ to reach the ‘spermatophoric organ’.
Within the spermatophoric organ the sperm are formed into a spiral mass and coated with the various membranes to become ‘spermatophores’ (sperm packets).
These spermatophores continue along another ciliated tube called the ‘vas efferens’ to be stored in the ‘spermatophoric sac’ or the ‘Needham’s sac’ until copulation.