Proteobacteria
Proteobacteria is the second largest group of bacteria.
This phylum contains 1,534 species – or 32.3% of all known bacteria.
Proteobacteria are all gram negative, but otherwise represent a diverse range of organisms such as the Purple Phototrophic, Nitrifying bacteria and Enteric bacteria – as well as the bacteria responsible for animal bioluminescence.
Here I will briefly introduce some of the better known groups.
Major Groups of Proteobacteria:
- Purple Phototrophic Bacteria
- Nitrifying bacteria
- Sulphur and Iron Oxidising Bacteria
- Spirilla
- Methanotrophs
- Acetic Acid Bacteria
- Free-living Aerobic Nitrogen Fixers
- Other Gram Negative Bacteria
- Enteric Bacteria
- Bioluminescent and related bacteria
- Rickettsias
- Hydrogen Oxidising Bacteria
- Sheathed Proteobacteria
- Budding and Stalked Bacteria
- Gliding Myxobacteria
- Sulphate and Sulphur Reducing Proteobacteria
Purple Phototrophic Bacteria
These are phototrophic bacteria – which means they get their energy from light.
Unlike green plants and cyanobacteria however, they do not give off oxygen in the process. This is described as anoxygenic photosynthesis. To do this they contain pigments very like chlorophyll called bacteriochlorophylls as well as a number of carotenoid pigments.
It is these carotenoid pigments that give Purple Phototrophic Bacteria their colours – purple, red and brown.
They are divided into two groups, depending whether or not they use hydrogen sulphide, H2S, as an electron donor for carbon dioxide reduction. If they do, then they are called Purple Sulphur Bacteria. If they don’t do this under normal conditions, then they are known as Purple Non-sulphur Bacteria.
Purple sulphur bacteria normally respire anaerobically and the presence of oxygen hinders their growth. Their favourite habitat is in the illuminated but anoxic zones of aquatic environments. Typical genera are Ectothiorhodospira and Chromatium. Some species are extremely halophilic (salt loving) and can be found in soda lakes and salterns.
In contrast to this group is the Purple Non-sulphur bacteria.
These were originally named because it was thought they could not use sulphide in their metabolism, though it is now known they can if necessary. However, the levels of sulphides utilized by Purple Sulphur Bacteria are as lethally toxic to Purple Non-sulphur bacteria as they are to most other species of bacteria.
A few species are capable of anaerobic growth, though most species are aerobic.
Most species can utilise a variety of carbon sources, from carbon dioxide through simple sugars to aromatic compounds and are also phototrophic, i.e., they derive their energy from light.
Like all groups in this section they are morphologically diverse (have very different shapes). They can be rods, curved rods, ovoid, flagellated, ring-shaped or spiral.
Typical genera are Rhodospirillum and Rhodopseudomonas with 15 species each and Rhodobacter with 8 known species.
Nitrifying Bacteria
These bacteria are linked by their ability to grown chemolithotrophically at the expense of reduced inorganic nitrogen compounds.
Most species are obligatory chemolithotrophs, the major exception being Nitrobacter which can grow on organic compounds and thus can live chemoorganotrophically.
These bacteria can be divided into two groups: those that oxidise ammonia to nitrite and those that oxidise the nitrite to nitrate. The first group are called nitrosifyers and the second group are the true nitrifying (nitrate producing) bacteria. No bacterium is known which reduces ammonia to nitrate entirely.
These bacteria were among the first to show that chemolithotropic life was possible. This was proven by a guy named Winogradsky in 1889. Winogradsky went on the following year (1890) to show that some bacteria were chemolithotrophic autotrophs. Meaning that they can grow in the absence of an organic carbon source, i.e., they fix carbon dioxide.
Previously, it was believed that only green plants or other organisms containing chlorophyll could turn inorganic carbon dioxide into organic carbon products.
Nitrifying bacteria are widespread in soil and aquatic environments, where they are an important part of the nitrogen cycle.
Important nitrosifying genera are Nitrosomonas, Nitrosococcus and Nitrosolobus. Important nitrifying genera are Nitrobacter, Nitrococcus and Nitrospira.
Sulphur and Iron Oxidising Bacteria
These organisms are primarily linked through their ability to reduce sulphur compounds, producing SO42- and 2H+ (which inevitably becomes H2SO4 = sulphuric acid) from H2S, S and S2O32- with the addition of either O2 or H2O.
One species, Thiobacillus ferrooxidans can also grow by oxidising ferrous iron such as iron pyrites, FeS2.
It does this in nature, where it is a blessing to miners because it makes the iron more available – but a curse otherwise because it causes environmental acidification when releasing other associated heavy metals into the environment. Thiobaccillus species are rod shaped bacteria.
Beggiatoa is a filamentous gliding bacterium which oxidises sulphur compounds in a similar way to Thiobaccillus. It is commonly found in sulphur springs, sewage works and hydrothermal vents.
It can also be found in rotting piles of seaweed and in the rhizosphere of plant roots – in swamps where it lives at the boundary between the aerobic or oxic layer around the roots and the anoxic soil nearby.
Here it is probably beneficial to the plants by oxidising hydrogen sulphide (H2S) which is highly toxic.
Thioploca, Thiotrix and Leucathrix are also filamentous sulphur-oxidising bacteria with the habit of aggregating within a star or rosette sort of shape, with their filaments united in a central core. These species are primarily found in marine habitats where they can form thick mats.
Hydrogen Oxidising Bacteria
Hydrogen oxidising bacteria are capable of growing with Hydrogen (H2) as an electron donor and oxygen (O2) as an electron acceptor.
Thus with the help of Nickel-containing hydrogenases they perform the reaction O2+2H2=2H2O.
This is known as the ‘Knallgas’ reaction and simply means that one oxygen molecule plus two hydrogen molecules – with the help of the bacterium – becomes two water molecules.
Nearly all the bacteria in this group are facultative chemolithotrophs, meaning that they can also grow chemoorganotrophically if they have to.
Some species in this group can grow autotrophically, using CO2 as a carbon source.
Others can use CO (Carbon monoxide) as an energy source.
Bacteria which oxidise CO to CO2 are known as carboxydotrophic bacteria. Carboxydotrophic bacteria such as Pseudomonas carboxydororans are important because CO is toxic and – although our cars have been releasing carbon monoxide into the atmosphere for years – the total levels have not risen.
This is believed to be as a result of the work of these bacteria in the soil.
Methanotrophs
These are methane oxidising bacteria (methane to methanol).
Methane is produced by other prokaryotes, particularly many Archaea. Methane’s oxidation to methanol is an important aspect of the carbon cycle.
Methane is often produced by anaerobic organisms while oxidation reactions need oxygen. Methanotrophs are therefore aerobic. They can often be found at the boundary layer between anoxic and oxic environments. Some Methanotrophs are also Methylotrophs, meaning that they can utilise other single carbon sources such as methanol, methylamine and formate (CH3OH, CH3NH2 and HCOO).
There are many bacteria however which are Methylotrophs, but not Methanotrophs, i.e., they cannot grow using methane. As far as is known, all methanotrophs are limited to using single carbon sources – thus they cannot utilise even simple sugars to grow.
Some marine mussels and sponges which live near hydrothermal vents – where methane is often released – have methanotrophs living symbiotically with them.
The bacteria get a safe home and a stable environment and the hosts get the excreted excess carbon compounds from the bacteria.
Note: The genus Zymonas contains two species which convert sugars to ethanol. These are used in many tropical areas of S. America, Central America and Asia to ferment alcoholic drinks. The alcoholic drink, ‘pulque’, made from the juice of the Agave plant is primarily fermented by Zymonas bacteria.
Acetic Acid Bacteria
These bacteria are the bane of amateur brewers because of their ability to turn a brewing wine into vinegar.
Basically, they partially oxidize various organic compounds (particularly ethanol) into acetic acid.
When not ruining wine these bacteria (Acetobacter spp) are used commercially to make vinegar.

They are also unusual because of their ability to synthesize cellulose (otherwise only known from plants), which they excrete as a matrix around themselves.
Free-living Aerobic Nitrogen Fixers
These are an ecologically important group of bacteria that live in the soil or in water.
The are united by their ability to fix nitrogen. This means that they can take gaseous nitrogen (N2) from the atmosphere and combine it with Carbon and Hydrogen, etc., to make organic molecules.
Nearly, if not all, the organic molecules in the world (i.e., all of you, your pets and me) are derived from bacterially fixed nitrogen.
Important genera in this group include Azotobacter, Azomonas, Azospirillum and Beyerinckia.
Other Gram Negative Bacteria
This is a group of related bacteria that don’t fit into any other group.
They contain both free-living species and species found living inside animals. Among this latter division is the genus Neisseria, including the species Neisseria gonorrhoeae which is responsible for the human sexually transmitted disease gonorrhoea.
Other species of Neisseria, plus species of Kingella, Moraxella and Acinetobacter may be pathogenic (disease-causing) at times.
Enteric Bacteria
The human gut (from mouth to anus) forms a wonderful habitat for bacteria.
It is a warm, moist, stable environment with plenty of food. This also applies to the other fully warm-blooded animals – i.e., mammals, birds. Therefore it is not surprising that their (and our) guts are home to huge numbers of bacteria. Human faeces generally comprise 30% (dry weight) of dead bacteria.
Many of the bacteria in this group are among the common inhabitants of warm guts.
Among them is the most studied of all bacteria, Escherichia coli. E. coli may be nutritionally beneficial to its hosts because it will release vitamins such as vitamin K.
E. coli comes in many strains some of which can be pathogenic, causing diarrhoea in children and urinary infections in older humans. It is, therefore, sometimes a mixed blessing.
A more vitriolic member of this group is Salmonella. Most members of this genus are pathogenic, causing diseases such as typhoid fever and gastro-enteritis in humans and other warm-blooded animals.
A third pathogenic member of this group is the genus Shigella.
Shigella dysenteriae causes a severe form of gastro-enteritis in humans called ‘gastric dysentery’. It can be transmitted both by water and through food.
Finally, the genera Proteus and Enterobacter can cause urinary infections in humans. While Klebsiella pneumoniae occasionally causes pneumonia. Most Klebsiella species can also fix Nitrogen.
The genus Serratia occurs naturally in soil and water as well as in the guts of insects and vertebrates.
Bioluminescent and Related Bacteria
The two key genera in this group are Vibrio and Photobacterium.
They are linked by being gram-negative, facultatively aerobic rods and curved rods with a fermentative metabolism.
Two species of Vibrio cause disease in humans. Both, like most Vibrio species, come from aquatic habitats. Vibrio cholorae causes cholera, one of the most common infectious diseases in third world countries. The other species, V. parahemolyticus, inhabits the guts of fishes and is believed to be the cause of numerous gastro-enteritis outbreaks – mainly in Japan, but also in other countries.
A few species of Vibrio and most species of Photobacterium possess the interesting property of emitting light.
The chemical reaction that results in the light being emitted is mediated by the enzyme luciferase and the reaction is an oxidation reaction.

Bioluminescent bacteria are mostly associated with fish. Either with live fish, where they live in the light organs of these fish, or with dead fish when they give them an eerie glow.
In live fish, the bacteria often live in spherical cells. As the light emitting reaction only occurs in the presence of oxygen, the fish can control the light flashes or glows by controlling the oxygen flow to the cells containing the bacteria.
Rickettsias
Rickettsias are an unusual group of bacteria, in that all but one species are obligate intracellular parasites.
So far, scientists have been unable to grow them in cultures outside of cells.
Nearly all species are pathogenic, causing diseases such as Typhus, Spotted Fever, French Fever, Q Fever and Ehrlichiosis in humans and Potomac Fever in horses.
Important genera include Rickettsia, Rochalimaea, Coxiella and Ehrlichia.
Spirilla
These are a group of roughly spirally shaped proteobacteria. They’re a diverse group showing a considerable range of spirallity.
Perhaps the most interesting genus in this group is Bdellovibrio. Bdellovibrio contains 4 species, all of which are predators on other bacteria.
They are all small bacteria, generally less than a tenth of the size of their prey. They attack by dissolving a hole in their prey’s cell wall, then living between the cell wall and the cell membrane and feeding on the cytoplasm.
Other noteworthy members of this group include Campylobacter and Helicobacter, which are pathogenic to humans – causing acute enteritis, chronic gastritis and peptic ulcers.
Ancylobacter, a ring-shaped bacterium and Magnetospirillum magnetobacterium, a curved rod shaped bacterium, contain 5 to 40 magnetic particles (Magnetite Fe3O4 or Greigite Fe3S4).
These are called magnetosomes and they allow the organism to align itself in relationship to magnetic fields. Cells in the southern hemisphere have the opposite polarity to cells in the northern hemisphere.
Scientists do not really understand the ecological value of this characteristic.
Sheathed Proteobacteria
This small group of chemoorganotrophic bacteria is unique in their habit of living within a sheath.
Because growth only occurs along one axis, this sheath eventually becomes a tube – with the bacteria at the ends secreting more sheath as division within the tube pushes them outwards.
When nutrients are low, the individual cells develop a flagella tuft at one end which allows them to disperse. These highly motile cells are called swarmers.
The various species in this group of proteobacteria are found in nutritionally rich waters such as sewage works. They will also form blooms in autumn, when fallen leaves settle thickly at the base of streams.
Budding and Stalked Bacteria
A variety of Proteobacteria produce extensions from their main cell.
These can be of various shapes, but are of smaller diameter than the mother cell. The extension of the cell wall and cell membrane contains cytoplasm. Such extensions are called Prosthecae (singular, Prostheca). These prosthecae are often involved in reproduction.
Two fundamental differences between this form of reproduction and normal binary fission are:
- After cell division the two cells are not of equal size – there is obviously a mother cell and a daughter cell. In binary fission both new cells are of equal size.
- The new cell wall that separates the cytoplasm into two sections starts from a single point. This is called ‘Polar Growth’. In most bacteria the new cell wall starts to form all across the cell. This is called ‘Intercalary Growth’.
Bacterial prosthecae are often simple hyphae which then form a bud at the distant end.
This bud grows to be a new daughter cell. In the case of some species such as Hyphomicrobium sp. the smaller daughter cell eventually develops a flagellum and swims away. When it is full-grown it loses the flagellum and starts to grow prosthecate hyphae of its own.
Some bacterium can have more than one hyphae at a time. As in normal binary fission, the nuclear material replicates before the dividing cell wall is built up.
Some bacteria produce a holdfast at one end of a hypha and use this to attach themselves to the substrate.
Such bacteria are called stalked bacteria because they appear to be resting at the top of a stalk. In some other bacteria the appearance of a stalk is given by a similar structure of extruded ferric oxide. This is in an organic matrix but not in form of a cytoplasmic filled prostheca.
An example of the latter is the genus Gallionella, while the genus Caulobacter produces ordinary stalked bacteria.
Gliding Myxobacteria
A few groups of proteobacteria can move by gliding along surfaces, often leaving a visible trail of slime.
By far the most interesting of these are the fruiting Myxobacteriaa. These are a group of rod shaped, gliding bacteria with the unusual habit of aggregating to form fruiting bodies when nutrients are low.
The cells come together and secrete a stalk – made mostly of slime – up which they then climb, to congregate at the top in a fruiting body.
The individuals then form myxospores, which can survive until it is environmentally suitable for them to live again.
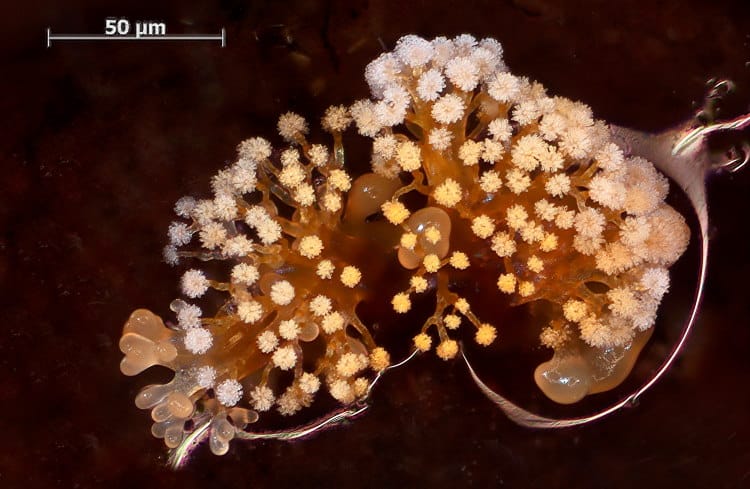
Example species are Mellitangium erectum, Chondromyces crocatus and Myxococcus stipitatus.
Finally, about 90 species of bacteria from over 20 genera are known which are obligate sulphur or sulphate reducers, living in anaerobic habitats.
Most species live in aquatic environments where the normal processes of microbial degradation of organic matter produce the oxygen free zones they require.
Desulfomaculum is more unusual in being a soil dwelling species and it is reasonably well known because it occasionally gets into tinned meat, where it results in a type of spoilage known as ‘sulphide stinker’.
A wide variety of sulphur compounds can be reduced by most species from elemental sulphur to thiosulphate and dimethylsulfoxide.
Species such as Desulfuromonas sp., which reduce elemental sulphur, often share their habitat with species of Green Sulphur Bacteria which oxidise H2S to S making a short complete cycle.
Other important genera include Desulfovibrio, Desulfobacter and Desulfobacterium.
Final Thoughts
Well, that’s all for no. I hope this brief introduction to the wonderful world of proteobacteria has been useful!