The Myxozoa; An Introduction
Taxonomically the position of the Myxozoa is still not entirely clear. Are they a class, a subphylum, or a superclass? However, regardless of the uncertainty over the exact level of taxonomic classification they should occupy over the last two decades it has become accepted that they are derived Cnidarians. Not only are they Cnidarians, but, with over 2200 known species, they are a very successful group of Cnidarians. In fact, some experts think there may be as many as 30,000 species waiting to be discovered.
The Myxozoa are all microscopic, endoparasites with complex life cycles that involve more than one host. As a group they have undergone extensive simplification of their body structures. Despite this they exhibit a bewildering complexity of lifestyles. Of the 2,200 species described so far only a couple of hundred have had their full life cycle described.
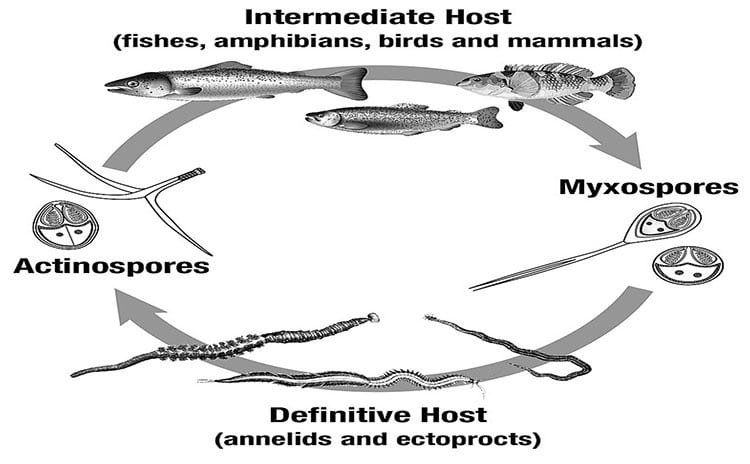
The Myxozoa are divided into two orders, which among other differences use different species groups as their definitive hosts. The majority of the Myxozoans occurr in the order/class Myxosporea. To date less than 20 species have been identified from the order/class Malacosporea. The Myxosporeans use mostly annelids and sipunculids as their definitive host and the Malacosporeans use Bryozoans. Both groups use fish, or occasionally other vertebrates, as their intermediate hosts.
Apart from DNA what the Myxozoa have in common with cnidarians is the possession of certain virtually identical, highly complex organelles. These are termed polar capsules in myxozoan spores where they serve for attachment to new hosts and they are the nematocysts in surface epithelia of cnidarians, serving for food capture.
Although Myxozoan spores are multicellular, the trophic body forms of almost all species are greatly simplified, reduced to syncytial plasmodia or to single cells.
General Biology of the Myxozoans
The Myxozoa are highly simplified animals. They have become simple through the evolutionary process of losing more complex anatomical features they once possessed but now no longer need. This process was involves the loss of the genes for these structures and most likely gradual. However, it has resulted in nearly all modern Myxozoans having secondarily lost all epithelial (skin or skin-like) structures. They also lack a nervous system, a gut, and cilia. Most lack muscles, though a few of the larger members of malacosporea retain muscles. Malacosporeans are also the exceptions in regard to epithelial structures, and are in some cases much larger than the more numerous Myxospreans, up to 3mm in length.
Myxozoans do not undergo any form of embryogenesis during development and no longer produce any true gametes. Reproduction occurs through the production of multicellular spores. The polar capsules which define these spores are not usually present in body cells.
Cell division does not usually involve binary fission, (the exceptions being in the Bryozoan phase of some Malacosporea). Instead cell production occurs through the use of endogeny, meaning new cells are produced within existing cells. After nuclear division, one of the nuclei and its surrounding cytoplasm become enveloped by endoplasmic reticulum, in this way a secondary cell is formed. This secondary cell lies in a membrane-bound vacuole in the primary cell.
Nuclear division by meiosis occurs in the invertebrate phase of both myxosporean and malacosporean cycles. However, this nuclear division is accomplished without the production of centrioles.
Recent research (January 10 2020) has shown that The myxozoan Henneguya salminicola (a parasite of Pacific Salmon), has no mitochondrial genome. This inevitably means it has lost the ability to perform aerobic cellular respiration (something previously thought basic to all Eukaryotes. This indicates that these core eukaryotic features are not ubiquitous among animals. The researchers analyses suggest that H. salminicola lost not only its mitochondrial genome but also nearly all nuclear genes involved in transcription and replication of the mitochondrial genome. In contrast, they identified many genes that encode proteins involved in other mitochondrial pathways and determined that genes involved in aerobic respiration or mitochondrial DNA replication were either absent or present only as pseudogenes.
The morphological simplification of the Myxozoa is reflected in the small sizes of their genomes (< 200 Mb), which are less than half that of free-living cnidarians, e.g. Nematostella vectensis (450 Mb),
Acropora digitifera (420 Mb) and Hydra vulgaris (1000 Mb). From “Myxozoans: Ancient metazoan parasites find a home in phylum Cnidaria”
Biology of the Myxosporea
The general life cycle for the majority of Myxosporeans is divided into a myxosporean phase in the vertebrate host and an actinosporean phase in the invertebrate host. Myxosporean spores
infect annelids and actinosporean spores infect fish. Myxosporeans may live inside the actual cells of the host’s body (intercellular), or they may occupy spaces in the host tissue in between individual cells (extracellular).
Growth of plasmodia follows an increase in the number of nuclei. Some of these nuclei are somatic (meaning they will become body cells), and some are generative (they will become the reproductive forms).
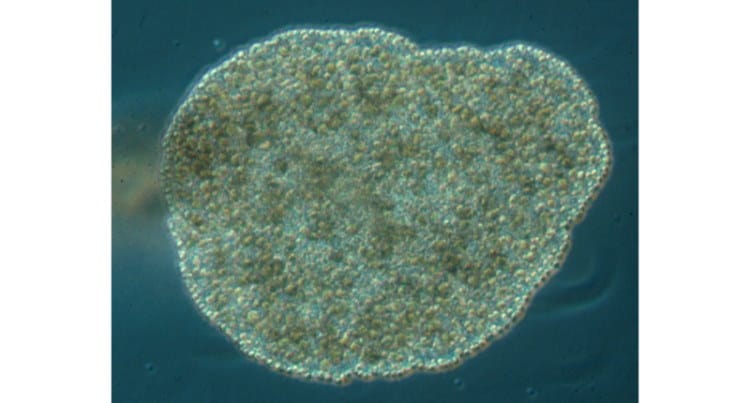
As a parasite that changes hosts as a natural part of its life cycle Myxozoans must have a trophic, or moving stage. In fact Myxozoans have two distinct and separate trophic stages.These extrasporogonic stages can take various forms. For instance some occur as pseudoplasmodia (uninucleate) while others exist as syncytial plasmodia (multinucleate). Alternatively they can be simply organized as a cellular layer around a syncytium or as hollow sacs or vermiform stages. Spores may be produced by either a plasmodium or by a pseudoplasmodium.
Plasmodia produce many spores (polysporic) and pseudoplasmodia normally produce two spores (disporic). Each sporogonic cell divides to give the requisite number of capsulogenic and valvogenic cells protecting one or two sporoplasms. In most genera spores contain a single binucleate sporoplasm but in a few cases (e.g. Sphaerospora & Unicapsula) there are two uninucleate sporoplasms. Morphogenesis of the polar capsules occurs in the capsulogenic cells. All cells in the myxosporean phase of myxozoans are believed to be diploid. This is the phase that occurs in the vertebrate host.
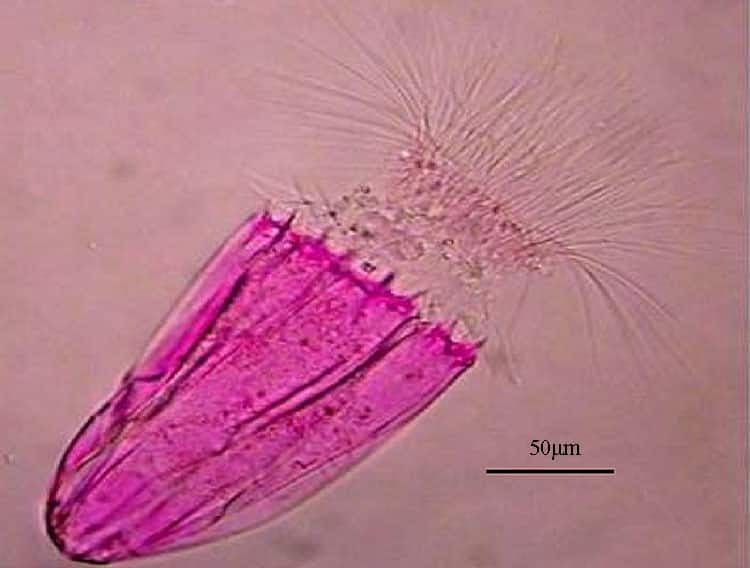
In the primary invertebate host, such as annelids the cycle of development is known as the actinosporean phase. After proliferation, a sexual cycle within pericytes culminates
in spore formation. The actinosporean spores (actinospores) differ from myxosporean spores in having a tri-radial symmetry (see diagram above) with three polar capsules and three valves, which often have elongated or bulbous extensions.
Biology of the Malacosporea
Unlike the Myxosporea, the Malacosporea parasitize freshwater bryozoans as their invertebrate hosts. They also have more complex trophic stages that develop epithelial tissues, have some muscle tissue and produce soft (uncuticularised) spores that are similar in the vertebrate and invertebrate phases. There are currently three described species: Buddenbrockia plumatellae, B. allmani and Tetracapsuloides bryosalmonae, all of which infect freshwater bryozoans as definitive hosts. Salmonids are intermediate fish hosts for T. bryosalmonae
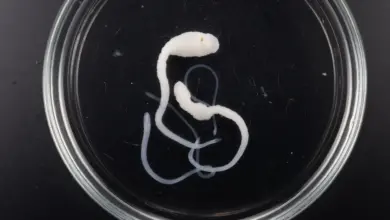
In B. allmani and T. bryosalmonae, the trophic stages within the bryozoan are saclike and exhibit an outer wall of epithelial cells joined by cell-cell junctions and underlain by a basal lamina . The species B. plumatellae is even more complex, with an additional internal epithelium that encompasses a central fluid-filled cavity and which, later in development, contributes to spore formation. Between these two epithelial layers and embedded in the extracellular matrix (ECM) are four longitudinal muscle blocks that enable the mature adults to undergo rhythmic sinusoidal or spiralling movements within the host body cavity. Nobody knows why they make this movement yet?
The early parasite stages are spherical in shape and attached to the host’s gut. These parasites grow in length, acquiring a more wormlike shape as they age. At an intermediate stage they can be found either attached or free-floating in the fluid-filled body cavity of the bryozoan host. The percentage of free-floating worms in a host increases as the animals grow older and reach later stages of development. However, even nearly mature worms may still be attached, though infrequently.
Late-stage and mature worms are distinguished by the occurrence of multicellular spherical spores in the inner cavity, but they differ little in size from the intermediate stage. In many cases, the cavity is not filled uniformly with spores. Rather the spores may be aggregated at one end, or in certain regions of the worm body. Mature worms are nearly always free-swimming in the host body cavity and exhibit vigorous movements. They are able to leave the host through ruptures or the coelom.
The muscles of the adult forms are arranged circumferentially in four layers that are situated between epidermis and inner epithelium. Each muscle block consists of two rows of obliquely oriented muscle cells.
Subphylum Myxozoa
- Class Malacosporea
- Order Malacovalvulida • 16+ living spp.
- Family Saccosporidae
- Order Malacovalvulida • 16+ living spp.
- Class Myxosporea • 2,200 + living spp.
- Order Bivalvulida • 707 living spp.
- Family Alatasporidae
- Family Auerbachiidae
- Family Ceratomyxidae
- Family Chloromyxidae
- Family Fabesporidae
- Family Myxidiidae
- Family Myxobilatidae
- Family Myxobolidae
- Family Myxosomatidae
- Family Ortholineidae
- Family Parvicapsulidae
- Family Sinuolineidae
- Family Sphaeromyxidae
- Family Sphaerosporidae
- Order Multivalvulida • 1,500 + living spp.
- Family Kudoidae
- Family Spinavaculidae
- Family Trilosporidae
- Order Bivalvulida • 707 living spp.
Image Credits:- Myxozoan Life Cycles by Paulo Selke – License CC BY-SA 4.0, Cover image from Myxozoa by Alexander Gruhl