Etymology = Latin Bis/Bi = two, and Valvae means leaves of the door – so – a two leaved door.
Species Number – Catalogue of Life: 2019 Annual Checklist = 9,115; Molluscabase = 12,535 (Marine – 11,141, Fresh Water – 1415)
Fossil Species :- Molluscabase:- 2,820
Condition of Typical Molluscan Characteristics
- Radula (a rasping “tongue” with chitinous teeth) = Absent
- Odontophore = (cartilage that supports the radula) = Absent
- Large Complex Metanephridia =
- Broad Muscular Foot = Present
- Large Digestive Ceca = Present
- Visceral Mass = Present
- Shell = Present (Two)
- Habitat = Aquatic – (Marine and Fresh water)
- Reproduction (Genders) = Commonly dioecious (having two separate genders) some Hermaphroditic (individuals are both male and female)
Introduction to the Bivalvia
Bivalves are a common group of animals. They occur in all aquatic habitats, however by far the majority of species are marine. Bivalves are known by a variety of common names, depending on species, or species group; Clams, Cockles, Fan Shells, Mussels, Oysters, Scallops and Shipworms are all species of bilvalves.
Bivalves are aquatic molluscs that possess two shells that are connected by a hinge. The two shells are commonly referred to as valves. These valves can be equal or unequal. Many, but not all, bivalves are bilaterally symmetrical. Bivalves possess no head but they do possess a large muscular foot that can often extend between the two valves. Mostly bivalves live within their shells, opening them only enough to circulate water and filter feed.
Although the Bivalves are the second most speciose group of molluscs today, after the gastropoda. There are more species known from the fossil record, 12,000+, than are currently alive.
Nearly all bivalves are edible, the exceptions being some tropical species of the genus Spondylus which accumulate saxitoxin, a powerful neurotoxin, (from the dinoflagellates they feed on). These species therefore become secondarily poisonous as a result of retaining the poison from their prey in their own bodies.
Bivalves are often confused with Brachiopods. This is because both groups have two shells and feed in a similar way, however they are not related. Brachiopods evolved earlier than bivalves and were very successful in the more ancient era. However after end of the Paleozoic era, 250 million years ago, the Bivalvia out competed the Brachiopoda and they are now far more numerous.
Bivalves can be extremely numerous where they occur, and they are widely spread throughout the marine ecosystem, especially the intertidal and sublittoral zones. Because they are diverse and common, bivalves are an extremely important part of the balance of many marine ecosystems. Bivalves are also important to humanity, mostly for food, but also for jewellry. More than 15 million tonnes of bivalves are harvested per year (average for the period 2010–2015). This is about 14% of the total marine production in the world.
Some Bivalve Record Holders
The Smallest:- The smallest know species of bivalve is Condylonucula maya, a minute marine clam which is less than 1 mm (0.04 inches) long as an adult.
The Largest:– The largest know bivalve is the Giant Clam, Tridacna gigas, which may be more than 137 centimetres (54 inches) in length and weigh 264 kilograms (582 pounds).
The Longest lived:- The longest lived bivalve know to science is the Ocean Quahog, Arctica islandica. This species is commonly reported to live over 100 years and in 2006 some scientists discovered a specimen which later turned out to be 507 years old, making it possibly the longest lived animal know to science. The story became popular in the press because the clam had to be killed to be aged, the specimen has been nick-named Ming in the common press.
The Fastest:- Bivalves are mostly sedantry or burrowing, however scallops are well know for swimming. But they are not particularly fast. Scientifically proven records are hard to find but the Australian Saucer Scallop Amusium balloti was clocked at 5.76 km/hr in 1989 by L. M. Joll
The Most Ancient:– The first fossil bivalves are found in the second half of the Tommotian period of the Cambrian era, meaning they lived about 525 million years ago. Three species are known from this time. Fordilla troyensis, Fordilla sibirica, and Bulluniella borealis.
The Deepest Living:– Bivalves occur commonly in coastal and near coastal water, but there are a few species in the genus Abyssogena which have evolved to survive the dark, the cold, and the pressure of the deepest oceans. The most abyssal of these is Abyssogena phaseoliformis which occurs at depths up to 6,400 meters (= 20,977 feet).

Basic Biology and Anatomy of the Bivalvia
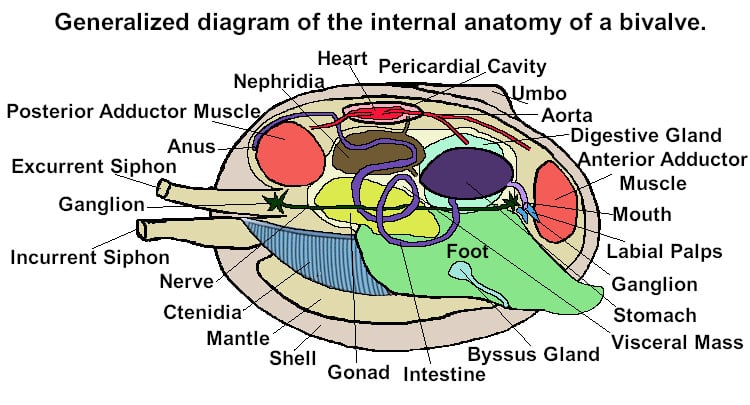
Bivalves have no head and no radula or odontophore. All bivalves possess a large muscular foot. Most bivalves have two tubes, or siphons, located toward the posterior end of the animal. These are called the incurrent siphon (which draws water into the mantle cavity) and the excurrent siphon (through which water and waste products are removed). The two siphons may be combined into a single siphon. They also have a basic three chambered heart, three or four pairs of ganglia, large (in most cases) ctendia (gills). Their digestive sytems comprises a pair of nephridia which extract biological wastes and a pair of gonads, a stomach, a pair of digestive diverticular and an intestine.
Feeding Ecology
Most bivalves are filter feeders however scientists believe the original bivalves were detritivores. The palaeotaxodonta have retained this ancestral habit. About 500 species of bivalves are carnivorous, mostly, as far as we know, catching and eating micro-crustations. These species have modified anatomies often involving reduced ctenidia, tentacled inhalant siphons and special stomach pouches to store and evict the shells of their prey.
Two other groups of bivalves with atypical feeding habits are the shipworms (family Teredinidae) and giant clams (family Tridacnidae). Shipworms are wood borers, they are capable of filtering food from the sea but they also digest the wood they burrow into.. Giant clams gain their nutrtion from symbiotic zooxanthellae (single celled algae) that live within their mantle tissue. The algae benefit from the dissolved waste products of the clam and the clams benefit from the waste products of the algae, this is called mutualism.
In the majority of bivalves water is drawn across the ciliated ctendia (modified gills) which are enlarged and folded. Food particles are caught in the filaments and swept down to the base where they become caught in a mucus stream that transports them to the labial palps and then into the mouth. Bivalves do not have a radula or an odontophore.
Digestion and Excretion
The basic configuration of the bivalve digestive system is similar to other molluscs. Food particles, once collected in various ways, are transferred to the stomach via the oesophagus. In filter feeders this involves an unusual organ called the crystalline style. This style is secreted by the style sac. Cilia within the style sac cause the style to rotate. This rotation serves to draw the mucus stream (with attached food particles) into the stomach. It also works to grind up material already in the stomach. As it turns the style dissolves into the stomach and releases digestive enzymes in the process.
Digestion of food particles occurs within the stomach and within the various digestive diverticula, sacks that open out from the stomach wall. Within the digestive diverticula the food is digested by macrophages. These are specialized cells that swallow a food particle and digest it within the cell, later releasing the digested material. This is called “intracellular digestion” to distinguish it from the more usual form of digestion involving a soup of chemicals, enzymes and food particles in the stomach and intestines which is known as “extracellular digestion”.
Many bivalves, especially fresh water ones separate the timing of their feeding (collecting food) and digestion. Meaning they actively digest food at a different time of day to when they actively collect food. This cycle of behaviour is often coordinated with environmental stimuli such as tides or day/night cycles.
Heart and Blood
Bivalves have a heart that lies within a membrane called a pericardium. The space the pericardium encloses is called the pericardial cavity. The bivalve heart is comprised of a central ventricle and two lateral auricles. As in other molluscs, the blood of bivalves is actually haemolymph. Oxygenated arises within the ctenidia (gills) and flows from them to the auricles.
From the auricles the blood is passed to the ventricle which pumps it out to the body along the anterior and posterior aortas. From these aortas the blood enters hemocoelic spaces in the mantle and visceral mass before returning to the heart via the ctenidia (gills) or the nephridia (kidney).
The majority of bivalves have no oxygen carrier molecule in their blood. However the most primitive bivalves do possess a haemoglobin. Meaning the loss of haemoglobin in most species is a characteristic that developed later in their evolution. This secondary loss of haemoglobin severely reduces the oxygen-carrying capacity of the blood. However this is compensated for by the large size of the ctendia (gills), the continuous rapid flow through of water, the relatively sedantry lifestyle of most bivalves and by the fact that oxygen can also be absorbed through the surface of the mantle.
Nerves and Brain
Bivalves have no head and no explicit brain. Instead their nervous system is comprised of a number of paired ganglia and a set of connecting nerve cords.
In the more primitive Palaeotaxodonta there are four pairs of ganglia, called cerebral, pleural, pedal, and visceral. The pedal ganglia are found at the base of the foot and the visceral ganglia are located below the posterior adductor muscle. The cerebral and pleural ganglia are located near the oesophagus.
In most bivalves the cerebral and pleural ganglia are fused together to make the cerebropleural ganglia. These ganglia are connected to both the pedal and the visceral ganglia by major nerve cords. In those bivlaves bivalves which have long siphons, there are additional siphonal ganglia.

Sensory Perception in Bivalves
How do bivalves sense the world around them? All bivalves have some sort of light sensing capability. In most species this is quite simple and merely allows them to sense gradients in light, meaning they can detect a shadow passing above them. This allows them to respond the presence of a potential predator by closing their valves, withdrawing their syphons and or burrowing down.
The eyes of bivalves can be as simple as just two cephalic eyes (connected by nerves to the cerebral ganglia) located on the ctenidia as in some members of the Arcoida. and Pterioida. Another type of eyes are ectopic pallial eyes, these are found on specialized folds of the mantle and also occur in Arcoida and Pterioida. Generally these are more numerous and are situated nearer the posterior end of the animal. They usually comprise a sensory pit containing light-sensitive, coloured, sensory cells.
More complex eyes called inverse pallial eyes occur in scallops and spiny oysters (The Pectinoidea). These are located on the mantle and are true eyes. They have a lens, a retina including sensory retinal cells, argentea (a silvery membrane) and a pigment cup. Simpler pallial eyes occur in the Cardioidea, in the Laternulidae, located on the inner margins of the mantle.
Bivalves also possess chemoreceptors and mechanoreceptors. Mechanoreceptors are sensitive to touch, in some cases these are located on short tentacles along the mantle margin. Chemoreceptor cells test the water quality, they are usually located near the siphons, but in some species, they fringe the entire mantle cavity.
Bivalves also possess an osphradium, which is also sensitive to water quality. In most bivalve the osphradium is found below the posterior adductor muscle. As well as this bivalves possess statocysts located at various places within the animal’s body. Statocysts allow the animal to sense its orientation, meaning it gives the animal a sense of balance. Each statocyst consists of a small sac-like structure lined with sensory cilia that detect the movement of a statolith. The statolith is a free moving mineral mass in the center of the sac. Movements of the animals body cause the statolith to bump into the cilia. Finally in the carnivorous Anomalodesmata, the incurrent siphon is surrounded by vibration-sensitive tentacles for detecting prey.

Musculature in the Bivalvia
The main muscular system in bivalves consists of the anterior and posterior adductor muscles. These are the strong muscles that allow the two valves to contract and the shell to close. They work in opposition to the ligament which acts to pull the valves apart. These muscles are contain two different types of muscle fibres. Striated muscle bundles are responsible for fast actions but are not capable of great strength, these allow the animal close its shell valves rapidly. Secondly there is a mass of smooth muscle bundles which work more slowly but generate great sustained strength and act to maintain the steady pressure necessary to keep the shell closed over a period of time.
In permanently sedentary bivalves, where one valve is always down, such as the oysters and scallops, and in the file shells that can swim by flapping their valves, only a single, central adductor muscle occurs. The anterior adductor muscle has been lost and the posterior muscle has moved to a more central position. In .
The numerous small mantle suspender muscles are what attach the mantle to the shell and they leave an arc-shaped scar on the inside of the valve called the pallial line (see below). Another important pair of muscles in a bivalves are the retractor muscles which allow the animal’s to move its foot. Some bivalves, such as oysters and most scallops, are unable to extend their foot and in these animals the retractor muscles muscles are absent. Other smaller pairs of muscles work to control the siphons and the byssus threads.
The Bivalve Ligament
Bivalves use muscles to close their shells, but they do not use muscles to open them. Instead they have a ligament. A mostly organic structure that acts like a kind of spring. The elastic nature of the ligament is what gives it its springiness. When the bivalve is shut, the ligament is compressed. As soon as the muscles relax the elastic nature of the ligament pushes the shell valves open again. This is why bivalve shells open when they die.
The typical ligament is comprised of two different types of layers. A wholly organic protein and collagen matrix layer (lamellar) and a semi-mineral (fibrous) layer . The semi mineral layer is a mix of aragonite strands and protein.
Simple hinges have only three layers; a single fibrous layer sandwiched between two lamellar layers. More complex ligaments have repeated layers of layers. Swimming bivalves such as scallops have more layered ligaments than more sedantry species.
The protein that is the basis of the ligament is called “abductin” and is unique to bivalves. It is also the only naturally occurring protein protein with compressible elasticity.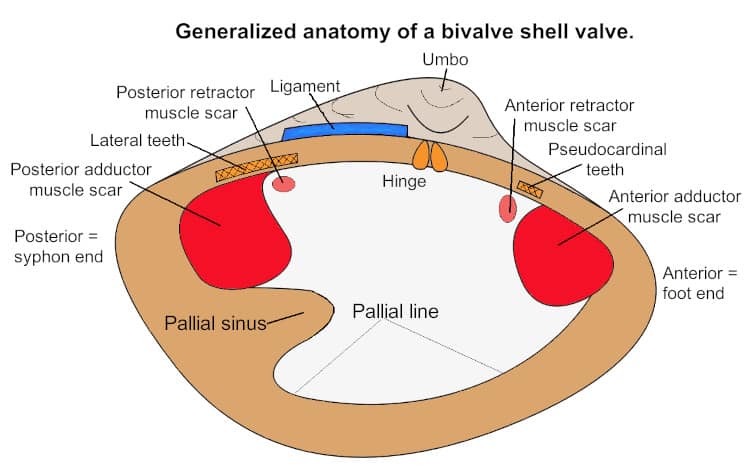
The Bivalve Shell
Bivalves possess a shell which is a part of their body and which acts as their exoskeleton. This shell is composed of two parts, a left side and a right side. Each shell valve also has an anterior and posterior edge. The anterior edge is where the foot protrudes beyond the valves and the posterior side is where the siphon is located.
The shell provides the animal with a home and with protection from predators. The physical nature of the shell is mostly two or more layers of calcium carbonate infused with an organic matrix. The Calcium carbonate layers can be all calcite, or some layers calcite and some layers aragonite. When aragonite is present it is often what gives some shells a pearly inner surface. The outermost layer of the shell is organic and is called the periostracum.
The bivalve shell is secreted by various parts of the mantle. The mantle itself is attached to the shell by a series of small muscles. On an empty shell these are seen as the pallial line which runs from the base of one adductor muscle to the other. The pallial sinus is an indentation in this line indicating where the syphons were positioned.
The umbo, and its tip which is called the peak, is the first part of the shell secreted. Not all bivalve shells have an umbo. In those that do the umbo often seems to point to one side or another, usually to the anterior side. So that when you hold the left hand/side shell with the inner surface towards you it looks like the diagram above.
When looking at the inner surface of a classical bivalve shell we can see certain marks, especially if the shell is fresh, if for instance you just had the animal as a part of your dinner. These marks represent various parts of the bivalve body, as detailed in the diagram above.
Bivalve shells can be bilaterally symmetrical, or not bilaterally symmetrical. They can posses an umbo or not possess an umbo. They can be smooth on the external surface or visibly ridged or adorned with spines. The position and number of the shell teeth varies considerable between groups and can even be absent as in the fresh-water clams in the genus Anodonta. . Finally the shells can be atypically shaped, forming a flattened cylindar as in the razor clams (family Pharidae), or greatly reduced and used only as a scraping tool as in the family Teredinidae.
Variations in shell morphology, are the backbone of bivalve taxonomy.

Reproductive Biology of the Bivalvia
The vast majority of bivalves are dioecious, meaning individuals are either male or female. Sexual dimorphism, where the males and females appear different, is rare. Hermaphroditism can take two different forms. Species can be either simultaneously hermaphroditic, meaning they have both male and female gonads at the same time, as in the Anomalodesmata. In this case self-fertilization is sometimes avoided by having either the male of female gonads mature in sequence, usually producing sperm first and ova later.
In consecutive hemaphroditism an individual is either male first and later becomes female (called protandry), or female first and later becomes male (called protogyny). The common shipworm, T. navalis, along with other members of the family Teredinidae, are consecutive hermaphrodites. The European oyster, Ostrea edulis, is a most unusual animal in that it changes it’s gender more than once, male, female, male female etc. This is called Rhythmical consecutive hermaphroditism.
Bivlaves have just one pair of gonads. In primitive bivalves these discharge their ova or sperm (or both) into the renal duct. However in most bivalves genetic cells are released through separate gonopores into the suprabranchial chamber.
Bivalve ova are generally small and many species synchronize their release of reproductive material. The spermatozoa of bivalves are unusual in that they have two tails. This synchronized spawning results in the release of large amounts of both types of gametes into the sea at the same time facilitating external fertilization. Hermaphrodites, and other brooding species which retain their ova within their mantle pallial cavity, usually bring in sperm from the surrounding water through their incurrent siphon.
Brooding and Larval Development
The embryos are then brooded within the modified folds of the ctenidia known as marsupia. There the fertilized eggs develop directly (without a larval stage), and the young are released as miniature adults. Although ctenidial incubation is most usual from of brooding, there are other patterns: egg capsules are produced by Turtonia minuta; the species Nucula delphinodontaa creates a brood chamber which is plastered to the shell of the female; and in members of the Carditidae the female shell itself is modified into a brood pouch. Finally, in a unique case, the 2mm long Grippina coronata broods it ova within its ovarian follicles and, thereby, provides the first example of intrafollicular fertilization and brooding in the Bivalvia, as of 2015.
For most marine species, however, the fertilized eggs float in the open water where they undergo indirect development. The first stage of this is called the trochophore larva. Trochophore larva are active swimmers. After a set amount of time the trochophore larva develop into veliger larva, which possesses a shell. These veliger use a ciliated velum for swimming. This velum can also be used for trapping minute particles of food. Following a period in the plankton, which varies from hours in some species to months in others, the veliger larva descend to the seafloor, where it metamorphosizes into small adults. During this process the the velum is lost, the foot develops and the ctenidia develop.
Ecology of the Bivalvia

Most bivalves live either partially or wholly buried in the substrate of the water body they live in, or firmly attached to a rocky or other solid substrate. Most species remain completely sedantry throughout their life, but others, such as scallops are much more capable of movement. Attachment to a solid substrate is either by means of secreted mineral-cement, or by byssus threads.
The group best known for using byssus threads are the mussels, both marine and freshwater species. The byssus threads are made of keratin and other proteins. It is secreted in liquid form that naturally creates a kind of foam. This foam is pumped and directed to the substrate by the mussel’s foot, each thread is about the width of a human hair. The mussel then adds another protein to the threads to make them sticky, resulting in an adhesive structure that is very strong.
Byssus is a remarkable because it is neither degraded nor deformed by water as synthetic adhesives are. This amazing quality has resulted in the genetics of byssus thread proteins receiving a lot of scientific study.
Oysters, thorny oysters and jingle shells form a permanent attachment to a hard substrate in a different manner. These are cementing bivalves. The cement themselves to the substrate using crystalline carbonate cements, basically the same material as their shell is made of.
Many permanently attached bivalves live in large or even massive congregations, particularly oysters and mussels. This habit makes farming them easy. The earliest oyster farms were developed in ancient Rome.
The vast majority of bivalves live buried in the substrate. This substrate can be mud, sand, shingle and small pebbles or even coral detritus. Bivalves are soft bodied, have no weapons and are not naturally toxic. The burying (hiding) lifestyle is essential for most species to survive as many other animals, including human beings are happy to eat them if they can.
The Unionida:- A Unique Lifestyle
The order Unionida are unique among bivalves and rather special in terms of the whole animal kingdom. The Unionida are all freshwater mussels (bivalves) and occur all over the world, with over 550 known species. As adults they are free living filter feeders like many other bivalves. However they possess a unique lifestyle in that they have larva that are obligate parasites. They are also one of the few groups of parasitic organisms in the world in which the larva alone are parasitic while the adults are free-living.
The Unionida, like many other bivalves, brood their fertilized ova in specialized ctendia called marsupia. Once they have finished developing the ova hatch into a specialized larva called a glochidium, pl. glochidia. In all the parasitic cases the glochidia are now released in the water to find a host.
For the vast majority of species this host is a fish, however at least one species, Simpsoniconcha ambigua, parasitizes a salamander, namely Necturus maculosus. Some groups of fishes, such as the centrarchids are more prone to being parastized than others. Also some species of fish seem to be popular with many Unionida. For example 17 different species of glochidia have been recorded from the White Crappie (Pomoxis annularis). Host specificity is generally low in the Unionida with some species, such as Pyganodon grandis parasitizing more than 30 species,
The glochidia larva attach to their host using either hooks or sticky mucous. However they seem unable to recognize suitable hosts until after they are attached. If they attach to an unsuitable host they later detach. Once attached to a suitable host the glochidium will encyst (embed in the host flesh and build a cyst around itself) within 36 hours, sometimes much more quickly than that. After encysting the glochidium feeds of the host tissues and develops into a juvenile mussel at which stage it will leave the host and sink to the substrate to begin living as a typical mussel.
The Unionida synchronize their reproductive cycles closely with the life cycles of their hosts. This is often important because adult mussels have sedantry life styles while many fish species move from one habitat to another throughout the year. Going up or down stream in rivers, from rivers to lakes or from lakes up into rivers and and from shallow to deeper water, or the other way around, in lakes. One advantage of this temporary parasitism of a mobile animal is that it helps the species to spread through, and between, habitats quickly.
What Eats Bivalves
Probably, on planet earth at this time the creature that eats the most bivalves is humanity. However much of what we eat is farmed.
In their natural environment, many creatures prey on bivalves. The Pacific walrus (Odobenus rosmarus divergens) is a major predator on bivalves in Arctic waters as marine clams are their favorite food item. Another mammal that feeds predominately on bivalves is the Sea Otter (Enhydra lutris). Sea Otters are famous for their use of rock tools to detach and break open tough clams.
Birds also prey on bivalves. Herring gulls are known to drop rocks onto mussels and oysters to break open their shells and Oyster Catchers feed almost exclusively on bivalves.
Fish also like to eat bivalves and a number of demersal species such as the common carp (Cyprinus carpio) regularly consume them. Another fish famous for its method of feeding on bivalves is the Harlequin Tuskfish (Choerodon fasciatus) made famous by David Attenborough’s Blue Planet ii video of one breaking open a small clam.
Under the water other invertebrates such as crabs, starfish, and octopuses all feed on bivalves. Furthermore more various species of predatory gastropods such as whelks have evolved to preferentially feed on bivalves. These fellow molluscs drill, or hammer, a hole into the bivalve’s shell and then suck out the flesh.

Taxonomy of Class Bivalvia.
The word clam is not a scientific or taxonomic word. It is a general-use word that is applied to a mixed group of bivalves that are commonly eaten.
Generally speaking, the taxonomy of the Bivalvia is a mess. The class Bivalvia is divided into two, five, six, or maybe seven subclasses; even when authorities agree on the number of subclasses they disagree on the names. The following scheme laid out by Newell in 1965 is still commonly used in many text books (with variations), however, it is not the scheme proposed by Molluscabase.
- Subclass Cryptodonta
- Order Solemyoida (Awning Clams)
- Subclass Pteriomorphia
- Order Arcoida (Ark Shells)
- Order Mytiloida (Marine mussels)
- Order Pterioida (Pearl oysters, pen shells, and fan shells)
- Order Limoida (File Shells)
- Order Ostreoida (Oysters and scallops)
- Subclass Palaeoheterodonta
- Order Trigonioida (Brooch Clams)
- Order Unionida (Freshwater mussels)
- Subclass Heterodonta
- Order Veneroida (Clams, cockles, razor shells)
- Order Myoida (Soft shell clams, Shipworms)
- Subclass Anomalodesmata
- Order Pholadomyoida
- Subclass Palaeotaxodonta
- Order Nuculoida (Nut Shells)
In 2010 the following publication reduced the subclasses of the bivalvia to two. Nomenclator of bivalve families; with a classification of bivalve families by R. Bieler, J. G. Carter & E. V. Coan

Bivalves and Humanity
Microplastic Pollution in Bivalves
The microplastic pollution of our oceans is now well documented. Microplastics are very small pieces of plastic that float in water, particularly in marine environments. Microplastics are defined as pieces of plastic either 1) less than 5mm in diameter, or 2) less than 1mm in diameter. Because most bivalves are filter feeders, because they have no way of distinguishing between microplastics and edible plankton, and because plastic is indigestible they accumulate microplastics in their bodies.
This accumulation of microplastics is passed on to and multiplied up in, the bodies of any animal that eats them. This includes human beings, who actually eat a lot of bivalves. Because microplastic contamination is such a new thing the effects it is likely to have on the human body have not been studied. However, studies on various smaller marine animals have indicated a number of negative effects. These have included greatly increased mortality, and less fatal negative effects on physiological functions such as respiration, nutrition, growth, and reproduction.
In a review published in 2018, Barbaroza et al listed over 100 species of marine animals that had been shown to have microplastics in their bodies. Of these 17 were bivalves including mussels, clams, and oysters. Furthermore, a study in 2014 estimated that European people who regularly eat mussels and oysters could be consuming as many as 11,000 microplastics per year.
As the amount of plastic in the oceans is increasing hugely every year these figures will rapidly become out-of-date with reality, but will remain useful as a baseline for future studies that will show an ever-increasing amount of plastic in our diets. For more on microplastics see the Earthlife page Microplastic World.
Commercial Value of Bivalves
Note:- The words clam, mussel, oyster and scallop are not species names but names for species groups. For example, the most commercially important edible oysters are found in the genus Crassostrea, i.e. C. angulata, C. gigas, C. rhizophorae, C. sikamea and C. virginica, however other genera of edible oysters include Magallana, Ostrea, Ostreola and Saccostrea.
Culinary Delights. Human beings love eating bivalves and between 2000 and 2010 commercial culture of some bivalves such as clams and cockles doubled. Bivalve production for human consumption is a massive industry. The total market value of marine bivalves is about 23 billion US$ per year (2010–2015). The largest producer of commercially farmed bivalves is China.
The total trade in bivalves in 2010 amounted to 14,616,172 tons. However, most of the bivalves consumed in the world today are farmed. A massive 12,913,199 tons of bivalves were produced by farming in 2010. The total world trade figures included 5,554,348 tons of clams, cockles, and ark shells; 1,901,314 tons of mussels; 4,592,529 tons of oysters and 2,567,981 tons of scallops.

Pearls. Pearl production is another important economic aspect of bivalve ecology. There are about 40 tons of cultured pearls produced each year. The major producers of cultured pearls include Japan, China, and Mexico. Various species of the family Pteriidae, particularly in the genus Pinctada produce marine pearls. Freshwater pearls come from species in the family Unionidae. Pearls occur naturally but most are cultured these days. A pearl can easily be identified as natural or cultured by examining its internal structure using x-ray radiography. Pearl oysters support a global cultured pearl industry with a value of almost $US 0.5 billion according to the book The Pearl Oyster published in 2011. The internal shell of the pearl oyster is called “mother of pearl” and this also has some commercial value.
Sea Silk. Sea silk is an extremely fascinating fabric. It is an exceptionally fine, rare, and ancient fabric that is made from the byssus threads of pen shells (in particular Pinna nobilis). Each thread is only about 6 cm long, but collected and spun, then treated with lemon juice they make an amazing golden-coloured thread that is finer than lepidopteran silk. Sea silk has been made since the times of ancient Greece. One of the reasons sea silk is so rare is that practically nobody is producing it anymore. In modern times it has never been successfully commercialized and is considered a dying culture.
Destructive Bivalves. Some species of bivalves are important agents in bioerosion, most notably by boring into calcium carbonate rocks and wood in the sea. Piddocks (family Pholadidae) bore into both concrete and wooden jetties weakening their structure. Shipworms (family Teredinidae) bore into wood and will attach themselves to ships with wooden hulls.
Marine mussels (family Mytilidae) settle on ships and reduce their potential speed by increasing the drag. They also colonize buoys, and wharves and may block seawater intakes into the cooling systems of power stations. In freshwater environments, the Zebra mussel (family Dreissenidae) can clog water-intake pipes and damage boats and bridges. They have been a problem in Europe since the 1800s. After being imported into the USA they upset the food web of the Great Lakes and threatened many native bivalve species with extinction. Causing millions of dollars in economic losses each year.
A few bivalves are host to human parasitic infections. Because bivalves are filter feeders they can collect anything that is in the water. Bivalves from waters that are contaminated with human made pollutants can and do accumulate these pollutants and potentially poison consumers. These pollutants can be disease vectors or chemicals and minerals. In 1978, Norwalk virus affected more than 2,000 people in Australia after they had eaten oysters. In 1988, an outbreak of hepatitis A arising from the consumption of clams (Anadara subcrenata) that were not properly cooked occurred in the Shanghai; China. It is estimated 290,000 people were infected and that 47 people died from the disease.